Driehoeksmossel - Dreissena polymorpha
Algemeen
Algemene kenmerken |
|
|---|---|
Naam soort(en)groep |
Driehoeksmossel - Dreissena polymorpha |
Regio |
Nederland, Europa |
Watersysteem |
meren, rivieren |
Natuurparameter |
weekdieren |
Factsheet opgemaakt door |
K.E. van de Wolfshaar |
Habitat beschrijving
Algemeen voorkomen
De Driehoeksmossel komt voor in rivieren, meren, plassen en kanalen en hecht zich aan allerlei hardsubstraat. Driehoeksmosselen zijn in de winter stapelvoedsel voor watervogels zoald de Kuifeend Aythya fuligula, de Tafeleend A. ferina, de Topper A. marila en de Meerkoet Fulica atra. Driehoeksmosselen zijn een doelsoort van zoete, grote stagnante wateren in het beneden rivierengebied.
Milieurandvoorwaarden
De milieucondities die van belang zijn voor de driehoeksmossel zijn zoet water, bodemtype, waterdiepte en watertemperatuur [#2], [#6].
Driehoeksmosselen vestigen zich op hard substraat zoals stenen en schelpen. Ze foerageren door algen te filteren uit het water. Hoge slibconcentraties in het water of op de bodem kunnen dit verhinderen en zorgen uiteindelijk voor verstikking. De diepte waarop mosselen voorkomen verschilt per gebied. Meestal komen ze voor op een diepte tussen de 2 en 5 m ([#2]). In ondieptes wordt de afswezigheid mogelijk veroorzaakt door de snelle temperatuurwisselen of door predatiedruk door mosselen of door een hoge wintersterfte als gevolg van een lange periode met zuurstofloosheid bij ijsbedekking ([#8]).
De groei en reproduktie van de driehoeksmosselen wordt beïnvloed door de watertemperatuur [#2]. In koude wateren (< 15 ˚C) is de periode van groei kort en is de reproduktie minimaal (Morton, 1969; Lewandowski, 1982; Strayer, 1991). Ook warme wateren hebben een negatief effect op de populatie-ontwikkeling (Walz, 1974). Sterftecijfers nemen toe bij watertemperaturen van > 26-32 ˚C (Strayer, 1991). Noordhuis en Houwing (2003) suggereren dat er mogelijk een negatief effect is van relatief warme winters. De eerste eieren en larven worden in de Poolse meren gesignaleerd bij watertemperaturen van 17-19 ˚C (Lewandowski, 1982) en in de Noord-Amerikaanse meren bij 13 ˚C (Hebert e.a., 1989).
In het IJsselmeer begint de jaarlijkse lengtegroei van de driehoeksmosselen in april en eindigt in september. Uit onderzoek van Reeders (1989) is gebleken, dat bij een watertemperatuur van 5-20 ˚C de filtratiesnelheid optimaal is. Beneden de 5 ˚C neemt de filtratiesnelheid abrupt af tot een geringe waarde en bij een watertemperatuur van meer dan 20 ˚C neemt de snelheid geleidelijk af. Volgens het onderzoek van Morton (1969) vindt alleen lengtegroei plaats bij een watertemperatuur van meer dan 11 ˚C en komt bij een watertemperatuur van meer dan 30 ˚C de lengtegroei geheel tot stilstand. De driehoeksmossel is zeer gevoelig voor snelle stijgingen van de watertemperatuur. Bij bestrijding van de driehoeksmosselen in koelwatersystemen wordt daarom de thermoshock-methode gehanteerd: bij een watertemperatuur van 35 ˚C wordt in een half uur alle aangroei gedood (Jenner en Mommen, 1985). De ontwikkelingsmogelijkheden van driehoeksmosselen in de Randmeren wordt beperkt door een aantal factoren, waaronder de snelle stijging van de watertemperatuur en grote dagelijkse temperatuurschommelingen die in het voorjaar in de ondiepe en beschutte meren kunnen optreden. Door deze veranderingen wordt de voortplanting van de driehoeksmosselen volledig verstoord (Vanhemelrijk e.a., 1993).
Beheer en ontwikkelingskansen
Met name veranderingen aan zoutgehalte, waterdiepte en watertemperatuur kunnen tot een verandering van het voorkomen van de driehoeksmossel leiden.
Dosis-effect relaties
Deze dosis-effect relaties zijn gebaseerd op een uitgebreide studie van TNO ([#2]) en een expert meeting ([#3]).
Er wordt onderscheid gemaakt voor meren en rivieren voor waterdiepte en stroomsnelheid [#2].
Stroomdiagram
graph[
rankdir=LR]
nodesep=0.5
"node0" [
label = "Zoutgehalte|Gemiddelde watertemperatuur juni tot augustus|Gemiddeld orhofosfaat|Gemiddeld zuurstofgehalte|Bodemtype|Slibgehalte"
shape = "record"
];
"node3" [
label = "HGI algemeen"
shape = "record"
];
"node0":f1-> "node3" [style=italic,label="minimum"]
[
id = 2
];
graph[
rankdir=LR]
nodesep=0.5
"node0" [
label = "Waterdiepte|Schelpen in bodem"
shape = "record"
];
"node3" [
label = "HGI meren extra"
shape = "record"
];
"node0":f1-> "node3" [style=italic,label="minimum"]
[
id = 2
];
graph[
rankdir=LR]
nodesep=0.5
"node0" [
label = "Waterdiepte|Gemiddelde stroomsnelheid oeverzone"
shape = "record"
];
"node3" [
label = "HGI rivieren extra"
shape = "record"
];
"node0":f1-> "node3" [style=italic,label="minimum"]
[
id = 2
];
Algemene dosis-effect relaties

zoutgehalte (g Cl/L) |
HGI |
|---|---|
0 |
1 |
1 |
1 |
1.1 |
0 |
5 |
0 |
Referentie: [#3]

temperatuur |
HGI |
|---|---|
0 |
0 |
13 |
1 |
17 |
1 |
20 |
0 |
25 |
0 |
Referentie: [#2]
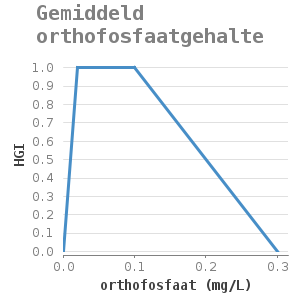
orthofosfaat (mg/L) |
HGI |
|---|---|
0 |
0 |
0.02 |
1 |
0.1 |
1 |
0.3 |
0 |
Referentie: [#3]

zuurstof (mg/L) |
HGI |
|---|---|
0 |
0 |
5 |
0 |
7 |
1 |
15 |
1 |
Referentie: [#2]

bodem |
HGI |
|---|---|
klei |
0.466 |
leem |
0.688 |
zand |
0.743 |
veen |
0.074 |
stortsteen |
0.879 |
Referentie: [#3]

slibgehalte |
HGI |
omschrijving |
|---|---|---|
0 - 10 |
0.753 |
slibarm |
10 - 25 |
0.45 |
matig slibrijk |
25 - 100 |
0.103 |
slibrijk |
Referentie: [#3]
Toegevoegde dosis-effect relaties voor meren

diepte (m) |
HGI |
|---|---|
0 - 2 |
0.287 |
2 - 5 |
0.76 |
5 - 10 |
0.634 |
10 - 25 |
0.205 |
25 - 50 |
0.033 |
> 50 |
0 |
Referentie: [#3]

Biovolume schelpen (ml/m2) |
HGI |
|---|---|
0 |
0 |
10 |
0.4 |
20 |
0.6 |
30 |
0.8 |
40 |
1 |
100 |
1 |
Referentie: [#7]
Toegevoegde dosis-effect relaties voor rivieren

diepte (m) |
HGI |
|---|---|
0 - 1 |
0.4 |
1 - 3 |
1 |
3 - 5 |
0.4 |
> 5 |
0 |
Referentie: [#2]

stroomsnelheid (m/s) |
HGI |
|---|---|
0 |
1 |
0.5 |
1 |
0.8 |
0 |
1 |
0 |
Referentie: [#2]
Onzekerheid en validatie
Deze dosis-effect relaties zijn gevalideerd voor het IJsselmeer ([#4]) en Markermeer ([#5], [#7]).
Toepasbaarheid
Deze dosis-effect relaties zijn toepasbaar op zoete wateren in Nederland.
Voorbeeld project
Volkerak-Zoommeer Habitat analyse ([#1]).
Referenties
1 Haasnoot, M. en Van de Wolfshaar, K.E.. Habitat analyse in het kader van de Planstudie/MER voor Krammer, Volkerak en Zoommeer. WL report Q4015. 2006
2 Duel, H. en Specken B.. Habitatmodel Driehoeksmossel en andere modellen voor het voorspellen van de populatie-omvang van de Driehoeksmossel Dreissena polymorpha in meren en rivieren. TNO-BSA werkdocument P94-026 1994
3 Van der Lee, G., H. Duel, S. Groot, H. Aarts, R. Pouwels (2000). Kwaliteit van het HEP-instrumentarium voor toepassing in het IJsselmeergebied. WL | Delft Hydraulics, WL-rapport T2391
4 Haasnoot, M. (2004) Memo validatie rekenregels Driehoeksmossel IJsselmeer.
5 Van de Lee, G., H.F.P. van den Boogaard, W.E. Penning (2001). Achtergronddocument voor aanvulling van het habitatinstrument: 1. Flexibilisering invoer; 2. Onzekerheidsanalyse; 3. Validatie. WL rapport Q3433
6 Noordhuis, R. & EJ. Houwing, 2003. Oorzaken en gevolgen van een vermoedelijke "crash" met betrekking tot waterkwaliteit, slibhuishouding en natuurwaarden. RIZA rapport 2003.016, Lelystad.
7 Van de Wolfshaar, K.E., M. Haasnoot, 2007. Toetsing dosis-effect relaties HABITAT. Pilot voor het Markermeer en de Voordelta. WL-report Q4327.
8 Strayer, D.L. (1991). Projected distribution of the zebra mussel. Dreissana polymorpha, in North America. Can. J. Fish. Aquati. Sci. 48:1389-1395.